Булдакова Т.И., Суятинов С.И.
Россия, Саратов, СГТУ
РЕКОНСТРУКЦИЯ ДИНАМИЧЕСКИХ СИСТЕМ НА НЕЙРОННЫХ СЕТЯХ
Рассмотрена задача моделирования методом реконструкции динамических систем по временным рядам. Отмечены недостатки существующих подходов. Показана возможность и преимущества решения подобной задачи на нейронных сетях. Особенности подхода исследованы на примере реконструкции системы по пульсограммам. Приведены результаты экспериментов на сети Хопфилда.
Многие проблемы управления, прогнозирования, распознавания образов связаны с задачей разработки математических моделей. Можно выделить два подхода к ее решению. Первый подход заключается в разработке модели, исходя из известных физических принципов и законов функционирования моделируемой системы. При втором подходе модель строят по временной реализации некоторой переменной состояния системы. В этом случае предполагается, что математическая модель представляет собой систему обыкновенных дифференциальных уравнений, приближенно описывающую эволюцию переменных состояния системы. Построение модели, или реконструкция динамической системы (ДС), собственно состоит в восстановлении модельных уравнений анализируемой системы, которые с заданной точностью способны воспроизводить экспериментально полученный временной ряд.
Остановимся более подробно на втором подходе, который наиболее применим к исследованию сложных систем, доступных для изучения только по своим реализациям. Наиболее ярким примером здесь являются медико-биологические системы, где достижения теории динамических систем могут находить свое практическое применение. В качестве примера рассмотрим задачу реконструкции динамической системы с целью диагностики и прогнозирования патологий по пульсограммам. Так, имея пульсовый сигнал (пульсограмму) и решая задачу реконструкции, можно оценить состояние здоровья человека, выявить патологии.
В данном случае интерес к пульсограммам обусловлен тремя причинами:
Существующие методы обработки пульсограмм основаны на определении характерных точек, изгибов, величин пиков, спектральном анализе. Однако мозг человека плохо приспособлен для выполнения арифметических операций. Мозг создает на своих нейронных сетях образы и оперирует ими. С достоверностью можно предположить, что, ощущая пальцами пульс, тибетские медики реконструировали в мозгу динамическую модель составляющей человеческого организма.
Следует отметить, что наблюдаемые временные реализации представляют такие установившиеся режимы, которые достигаются в пределе при t ® ? . Это подразумевает наличие в фазовом пространстве системы притягивающего предельного множества точек - аттрактора. Поэтому задача реконструкции подразумевает задание подходящего фазового пространства и формирование предельных множеств, соответствующих исходной временной реализации. Этой задаче посвящено достаточно большое количество публикаций (например, [ 1] и библиография в ней).
Изложенные в [ 1] подходы к проблеме реконструкции ДС характеризуются следующими основными недостатками:
Нейронные сети являются более гибким аппаратом моделирования [2]. Аппроксимация нелинейностей осуществляется нейронными сетями. Каждый нейрон сети реализует простую нелинейную функцию одного аргумента j ( y). Нейрон имеет несколько входов, на которые поступает вектор сигналов X. Значимость входов одного нейрона определяется вектором весов w. Для сети из n нейронов, на входы которых подается вектор сигналов X, будем иметь n векторов w, которые образуют матрицу весовых связей W.
Аргументом нелинейной функции нейрона является скалярное произведение (X,W), т.е. j (y) = j [(X,W)]. Здесь j - это сигмоидная функция активации нейрона, j = 1 ¤ ( 1 + e-y) . Результат преобразований передается на входы других нейронов. Нелинейное преобразование исходного образа X={x1,x2,…,xn} происходит по правилу
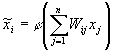 Нейронная
сеть представляет суперпозиции простых функций
одной переменной и их линейные комбинации и
производит аппроксимации нелинейных функций [ 3] . Так же, как и в
случае с полиномами, матрица W ищется
исходя из условия минимизации ошибки
аппроксимации. Главное отличие заключается в
том, что структура матрицы W (симметрия и
знаки элементов) задается первоначально
произвольно. На последующих шагах аппроксимации
значения элементов определяются, например,
методом наименьших квадратов. Избыточность
информации в сетевых связях обеспечивает
грубость аппроксимации при высокой точности.
Нейронная
сеть представляет суперпозиции простых функций
одной переменной и их линейные комбинации и
производит аппроксимации нелинейных функций [ 3] . Так же, как и в
случае с полиномами, матрица W ищется
исходя из условия минимизации ошибки
аппроксимации. Главное отличие заключается в
том, что структура матрицы W (симметрия и
знаки элементов) задается первоначально
произвольно. На последующих шагах аппроксимации
значения элементов определяются, например,
методом наименьших квадратов. Избыточность
информации в сетевых связях обеспечивает
грубость аппроксимации при высокой точности.
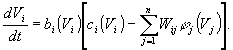 Помимо
аппроксимации нелинейностей нейронные сети
являются универсальным средством представления
оператора эволюции. В этом случае правила
преобразования образа (1) заменяются системой
нелинейных дифференциальных уравнений
нейронной динамики
Помимо
аппроксимации нелинейностей нейронные сети
являются универсальным средством представления
оператора эволюции. В этом случае правила
преобразования образа (1) заменяются системой
нелинейных дифференциальных уравнений
нейронной динамики
Здесь Vi(t) - состояние i - го нейрона в момент времени t; bi -усиление; матрица Wij и функция j имеют тот же смысл, что и раньше. В зависимости от выбора коэффициентов bi, ci получаются сети разных типов.
Для динамических систем подобного типа теорема Коэна-Гроссберга гарантирует существование областей притяжения - аттракторов. Эти области формируются в процессе синтеза матрицы W на основе исходных или эталонных образов. Эволюция состояния реконструированной нейронной сети приводит ее в область или точку равновесия. Эта область характеризуется локальным минимумом энергии, координаты которого соответствуют эталонному образу.
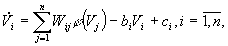 Из
уравнения (2) можно получить следующее
Из
уравнения (2) можно получить следующее
где bi - коэффициенты обратной связи, ci - внешнее воздействие на i - й нейрон. Полученное уравнение описывает сеть Хопфилда. Эта сеть выбрана для реконструкции динамической системы по данным пульсограмм.
Сети Хопфилда используются для запоминания статических образов, которые представляют точки аттракторов в фазовом пространстве нейронной системы.
Временные реализации реконструируемых систем представляют предельные множества другого типа. В простейшем случае - это предельный цикл. Возможны также более сложные образования. Поэтому особенность реконструкции на сетях Хопфилда заключается в формировании не точки равновесия, а предельного цикла. Эта особенность налагает дополнительные условия на правила синтеза матрицы W [4]. В частности, в экспериментальных исследованиях матрица была выбрана несимметричной. Помимо этого, структура сети учитывала то, что исходный сигнал является почти периодическим. Количество нейронов определялось из выражения n = (T/t )+1, где T - период исходной реализации, t - дискретность съема информации.
![]() Были
проведены две серии экспериментов. Первая серия
экспериментов базировалась на теореме Такенса [ 5] . В качестве
эталонных образов выбирались исходная
реализация вида
Были
проведены две серии экспериментов. Первая серия
экспериментов базировалась на теореме Такенса [ 5] . В качестве
эталонных образов выбирались исходная
реализация вида
и ее модификации, полученные простой перенумеровкой ai = ai+1. Таким образом, сети было представлено n эталонов.
Вторая серия экспериментов основывалась на последовательном дифференцировании исходной временной последовательности. В качестве фазовых переменных использовались значения a(t), a'(t) и a''(t). Поэтому сеть содержала 3n элементов. Выбор трехмерного пространства обусловлен тем, что оценка размерности Хаусдорфа оказалась равна 2,75.
Экспериментально доказана возможность реконструкции ДС на нейронных сетях в обеих сериях экспериментов. Решения реконструированных уравнений модели качественно повторяют основные особенности исходного сигнала, однако сходство в деталях проявляются не всегда. Кроме того, предельные циклы возникали только при определенных численных значениях исходных реализаций. Отметим, что вторая серия экспериментов менее чувствительна к числам.
Дальнейшие исследования необходимо направить на совершенствование методики определения матрицы связи и структуры сети, которые гарантированно приводили к предельным циклам.
Литература
| Site of Information
Technologies Designed by inftech@webservis.ru. |
|